
DuoBody平台介绍
DuoBody是genmab的双特异性抗体技术平台。这几年连续有三个基于DuoBody的双特异性抗体上市,包括:Amivantamab、Teclistamab和Talquetamab,这每一个都是如雷贯耳呐。之前可能很多人都不太想了解,但是随着这些双抗的上市大概率会碰到使用DuoBody的对照双抗从而导致大伙不得不去了解这个平台以及使用这个平台制作一些样品。除此之外DuoBody也是一个高通量的双特异性抗体筛选平台,可以让你在筛选双抗的时候事半功倍。
DuoBody简介
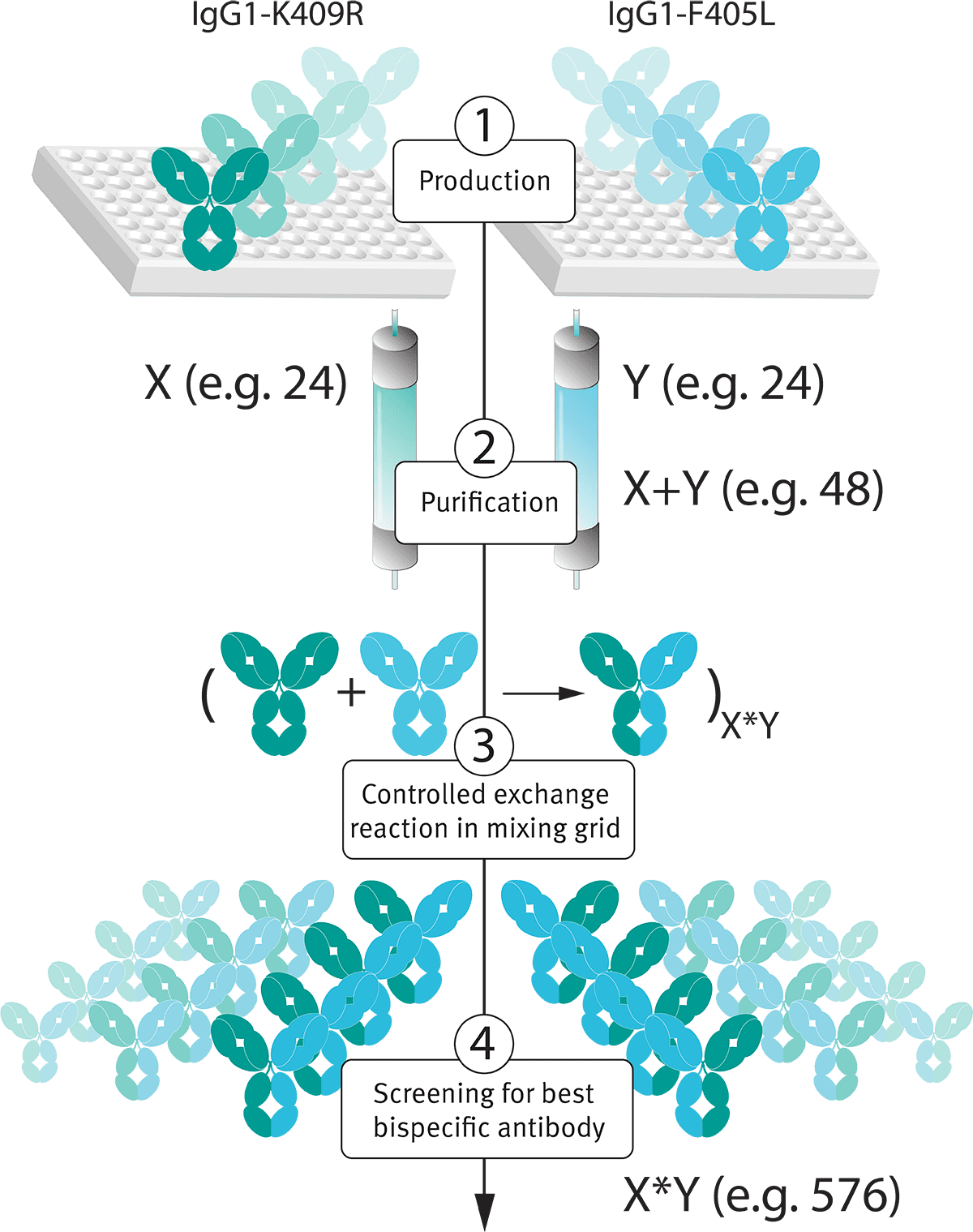
DuoBody的操作说起来很简单,首先分别在两个IgG1抗体恒定区引入L409R和F405L突变,将表达出来的抗体进行纯化,再经过“controlled Fab-arm exchange”(cFAE)就可以组装成双特异性抗体。有感兴趣的可以自己去关注下[引文1],genmab手把手教你怎么做。

DuoBody平台除了是双特异性抗体平台,同时也是一个高通量的双特异性抗体筛选平台,amivantamab就是使用这个技术筛选出来的。使用DuoBody平台可以快速组装双抗,而不需要一个个单独将双抗表达出来,你说目前有什么更加简单靠谱的技术让你在一个细胞里面表达类似的双抗结构呢?
IgG4与FAE的关系
IgG4抗体与其他IgG不同的特征之一是可以形成半个分子,并且在体内形成天然的双特异性抗体,这个过程叫“Fab-arm exchange”(FAE),除了人的IgG4,其他种属也存在类似的过程。
我们知道FAE主要由IgG4的铰链区控制,很多治疗性IgG4抗体都会携带S228P突变,就是为了消除这个过程。但是根据对IgG4恒定区的研究发现,除了铰链区以外CH3对FAE也有一定的促进所用,对IgG4的CH3进行一些突变也可以消除FAE,如409位点。通过对人IgG1恒定区CH3的研究发现K409的侧链刚好处于另一个CH3上的空腔处,这个空腔由L368‘、 K370‘、D399‘、F405‘和 Y407‘组成,K409与D399‘以及一个水分子形成氢键,其中水分子刚好可以阻止K409与K370‘发生电荷排斥,这个水分子同时也与S364和T411形成氢键。在IgG4中,R409的胍基与D399‘的侧链发生电荷相互作用,不过由于R的侧链体积较大,占了水分子的位置因此也缺失了水分子与S364、T411和 K370‘之间的氢键,从而导致R409和K370‘发生直接的电荷排斥。

在其他种属种也发现有FAE,不过从序列比对的情况来看这些种属发生FAE的机制与人IgG分析获得的结果并没有特别的相关性。具体到恒河猴中,通过与人IgG1序列对比发现CH3部分主要就3个氨基酸与人IgG有差异。

将rhChIgG4上的三个位点I350、T370和L405引如携带P228S的人IgG1中,发现只有F405L和三重突变体ITL可以发生Fab-arm exchange,因此在恒河猴IgG4的CH3中促进Fab-arm exchange的氨基酸为L405。

CH3之间的相互作用与FAE的关系
FAE的形成需要CH3之间的解离和结合,因此CH3之间的结合强度可能会影响这一过程。从下图的结果也能够看出对CH3部分的氨基酸进行突变会影响CH3之间的相互作用。

这边可以看到F405L突变可以降低IgG1 CH3之间的相互作用。通过结构分析可以得知F405原本处于T394’、K392’以及K409’形成的疏水腔中,当F突变成L后会增加CH3二聚体之间的空腔,从而减少CH3之间的疏水相互作用,使整体结构不再稳定。

且CH3之间的相互作用越低,发生FAE的程度越高。(C) IgG4-derived constructs,(D) IgG1-CPSC–derived constructs,5.3x10^-8^M似乎是一道门坎。

DuoBody设计
野生型的IgG1在适当的条件下也可以发生FAE,只是发生的比例比较少。因为FAE的发生需要两个步骤:首先需要打开重链间的二硫键,IgG4上S228使链间二硫键对还原条件更加敏感;其次CH3之间非共价结合的解开是发生FAE的限速步骤,这个过程依赖于CH3之间交界面上的氨基酸组成,之前提到的K409R或者F405L会减弱CH3之间的相互作用。因为正常情况下CH3的相互作用是没有特异性的,使用相同的CH3突变很难获得高比例的双特异性抗体,根据概率推算约为50%(前面图中的值是与IgG4对照的相对比值),因此需要设计出突变能够降低CH3同源二聚的相互作用让抗体能够发生FAE,但是异源二聚的CH3相互作用要足够强不仅可以在体系中更快发生异源二聚而且可以保持稳定避免继续发生FAE。

为了找到这种突变,对R409附近的氨基酸进行饱和突变(除了C和P),突变的位点为Y407、L368、F405、K370、D399,突变后与R409通过FAE组合成双特异性抗体,选择IgG1-K409R 和 IgG1-F405L的组合作为对照。可以看到有很多的组合都能形成双抗,并且验证了之前的设想。后续使用IgG1-K409R 和 IgG1-F405L进行进一步研究。


影响cFAE的因素
相同摩尔比例的IgG1-F405L-EGFR 和 IgG1-K409R-CD20使用25nM的2-MEA在不同温度下孵育(0,15,25和37 °C),可以看出随着温度的上升反应速度提高。37 °C孵育60min即可达到最高双抗浓度。锁定反应的温度37 °C和孵育时间90min调整2-MEA的浓度,可以看出随着2-MEA的浓度上升也能加快反应的速度,最低使用剂量为25nM,使用野生型IgG4获得的双特异性抗体比例较低。双抗的比例是使用质谱分析的结果,从这边也能看出野生型的IgG4生成双抗的比例最高为50%左右。

使用质谱以及CIEX可以分析出通过cFAE获得的双特异性抗体比例能够达到94~98%,而且普适性很广!

工艺放大
cFEA的工艺从小试到生产的放大基本是线性的,双特异性抗体的得率基本在93.0%-94.7%不等。

产物的纯度也与小试级别相当。

稳定性实验的结果显示,双抗在5 °C 6个月的放样过程保持稳定,但是在25 °C和40 °C会有降解。

对抗体恒定区功能的影响
突变位点位于CH3的内部,远离发挥效应功能的位点,因此这些位点的突变不会对IgG1的效应功能以及与FcRn的结合产生影响。从结果来看双抗能够保持野生型单抗的CDC(A、B)和ADCC(C、D),并且对双抗的PK也没有影响(E、F)。

讨论
还有其他临床前的体内抑瘤实验我就不在此多说了,毕竟Doubody平台已经通过临床的考验。Doubody平台不但解决了重链异源二聚的问题也解决了重轻链之间的配对问题,对抗体序列的突变更少,操作也特别简单,同时相对于其他双抗平台来说也能够做双抗的高通量筛选,这个优势是其他的双抗平台所不具有的。总结来说当年刚看到这个平台的时候很多人的第一反应就是一个项目要做两套细胞株工艺也更加复杂,但是基于Doubody的双特异性抗体上市似乎也说明这个平台确实有过人之处。从强生的两个CD3双抗Teclistamab和Talquetamab来看使用的CD3序列就是一样的,不晓得具体工艺是怎么干的,是不是3个细胞株就可以拼成两个双抗。
参考文献:
-
Controlled Fab-arm exchange for the generation of stable bispecific IgG1
-
Species-Specific Determinants in the IgG CH3 Domain Enable Fab-Arm Exchange by Affecting the Noncovalent CH3–CH3 Interaction Strength
-
Efficient generation of stable bispecific IgG1 by controlled Fab-arm exchange
-
Discovery of amivantamab (JNJ-61186372), a bispecific antibody targeting EGFR and MET
